从事物理学研究并涉及生物学边缘的人,对物理学原理与生物现象之间的相互影响有了初步的概念。应该说,对生命的研究应被视为现代物理学中令人激动的、同时也是真实存在的一部分。

盘桓在生物学和物理学之间的难题给物理学家挑战生物学提供了一个难得的机会;同样重要的是在这一过程中,物理学家通过新方法思考问题的同时,也丰富了物理学学科。
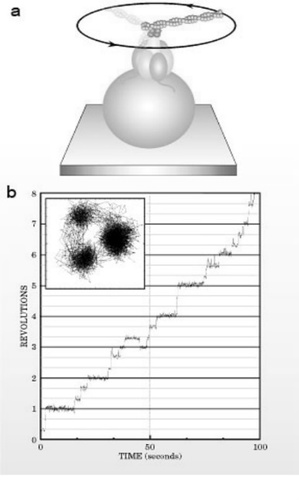
图1. 难以置信的小ATP合成酶发动机向腺苷二磷酸添加磷酸盐,产生三磷酸腺苷。当逆转时,它把存储ATP的化学能转化为旋转机械能。(a)添加到ATP合成酶轴上的肌(动蛋白)丝使轴的旋转在光学显微镜下被显示出来。每个三磷酸腺苷转化为二磷酸腺苷反应导致肌动蛋白推进器逆时针旋转120度。在此实验中,ATP合成酶的亚聚体附加到橙色的线上,结构显示推进器位置离散的变化陪伴着化学反应。
保罗 · 狄拉克(Paul Dirac)在1931年发表的那篇经典的磁单极子论文时谈到:目前理论物理学界存在的一些基本难题有待解决,如涉及量子力学方面的相对论方程和原子核的本质(更多的难题紧随其后,包括生命的本质问题等),这些问题的解决也许要对一些基本概念进行修改,这种修改比以前的任何一次都要来的彻底。
狄拉克认为,生命的本质不仅是生物学家的基本问题,同样也是物理学家的中心问题。然而,他的观点当时并未被沉浸在量子论革命带来的兴奋中的人们广为接受,物理学界只有一小部分人站出来面对他提出的挑战。
今天,人类正在收获近一百年来对量子论世界进行探索而取得的成果。许多重大问题已经得到解决。随之,人们正在探索如何将量子论应用于各种工程技术领域,如通讯、度量衡和计算。由此人们会问,如果这些占据了物理学主流地位的基本理论业已成熟的应用于工程实践,那么下一步物理学家需要研究的重大课题是什么?
我们认为,物理学家面临的重大挑战应该包括如何解释这个令人困惑、千奇百怪的——从可以自我复制的分子到大吃特吃的细菌,再到有意识的思维和行动的生物体——纷繁复杂的生命世界。
此外,分子和结构生物学、生物化学和遗传学取得的令人兴奋的成就引发了生物学信息的大爆炸,包括基因的表达及表达量是何时、何地联系在一起的。类似的信息是以图像的形式报告细胞的张力、速度和曲线方面的数据。尽管场被认为是一门“软”科学,但物理学家常用的定量分析现在几乎能熟练地应用于所有的生物体。就如天文学上采用光谱测定法进行天文观测所面临的巨大机遇一样。
对物理学家而言,生命意味着许多有趣的问题。我们在讨论以下三个物理学和生物学之间相交叉的问题时,往往将物理学作为综合思考的基础。首先,在描述组成生命基础的分子机器时,那些分子机器的运作为什么是如此的令人着迷,就是因为它们存在于某一结构内。在那儿,能量和长度曲线将不同的现象汇集到了一起。第二,是与生物学的“多体”问题有关,特别是细胞内的大分子组装的协同活动。第三,关注生物动力学的理论需要,不仅是生物体的多体特征,还考虑到它们远离平衡系统的运作。
我们这些从事物理学研究并涉及生物学边缘的人,仅仅对物理学原理与生物现象之间的相互影响有了初步的概念。应该说,对生命的研究应被视为现代物理学中令人激动的、同时也是真实存在的一部分。
生命机器
分子机器是生命的基础。DNA(脱氧核糖核酸),一种编码产生生物体的大分子也许是生命的信息存储器,但是它需要一组将信息阅读和翻译直至发挥作用的机器。细胞的毫微米大小的机器主要是蛋白质分子,虽然有小部分来自RNA(核糖核酸),它们能进行令人吃惊的复杂操作。几乎细胞内所有的重要任务都由它们执行:新陈代谢、繁殖、应对环境改变等等。它们的复杂程度令人难以相信,可以说达到了纳米技术的顶峰。然而,目前对它们的组装或作用的广义理论都还没有(理解单一的物理学基本原理很容易),所缺的是将各种现象升华并结合在一定框架内的非凡预见力。
蛋白质与二原子分子大相径庭,它反映了物理学和化学的传统边界。由于多种原因使量子力学在它们身上的作用可以忽略不计。当然,如果是发生在酶活化过程中,最终必须求助量子力学。在对日常的物体进行动力学描述时,量子力学同样会被忽略:在最小的长度标尺上,所有原子完全遵循量子论,并不需要普朗克常数来表达和应用最小作用原理。事实上,如果要求完全用量子力学来解释多种物理现象,其范围从蛋白质行为到星系临界现象是很难做到的。而作为分子聚合体的蛋白质,却经常被人们用来进行综合统计力学的分析。换言之,它们在本质上被作为经典的研究对象。
一个分子能发挥多大作用?想想ATP(三磷酸腺苷)合成酶这个例子吧。这种大分子的组装是细胞工厂制造ATP不可或缺的部分。我们不准备深入探讨细胞中ATP合成酶的生物学作用,仅考虑在隔离状态下它能做什么?它是一台回旋发动机。由于存在质子梯度,这台发动机在添加磷酸盐基到二磷酸腺苷上产生ATP时,触发了它的轴的转动。每天,细胞通过这种磷酸盐反应来制造与你体重大体相当的ATP分子。
但是这并非全部:ATP合成酶还能反向运转消耗ATP,伴随ATP分子的水解,ATP合成酶的中轴翻转120度,直接把化学能转化为机械能。通过一系列将分子驱动器粘附到轴上的试验,再通过光学显微镜取像,就能清晰地展现这种反向运转(见图1)。ATP中的驱动器旋转,使热力学功效达到90%。尽管纳米技术取得了巨大进展,然而具有类似功效的无机材料尚未被制造出来。另外,如ATP合成酶之类的分子机器将化学能转化为机械力的基本原理,仍有许多问题有待解决。
在嘈杂的环境中工作
分子机器运行在一个能量和长度相同、但加工过程不同的主机上,除了令人好奇以外,还增添了对细胞机器进行分析这一责任。图2显示了热能、化学能、机械能和静电能是怎样与相应大小的对象相匹配的,同时也表明了能量的汇合过程。当特定大小的生物大分子相互靠近时,所有能量聚集于一点。由于从亚原子大小到肉眼可见之间存在20多种大小不同的能量,因此这种聚集非同寻常。一般来讲,当热能在确定其他性能之间的相互影响丰富了分子机器的活动。例如,热能效应允许散射、构象变化、氢键分离等过程,其过程连续用大分子复制和阅读DNA进而发挥作用,为肌肉(如肌浆球蛋白)提供能量。
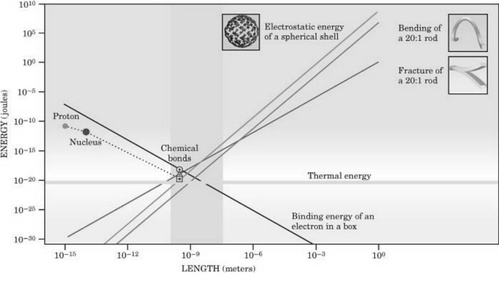
图2. 能量的汇合在该图得到显示,热能、化学能、机械能,还有随物体规模而定的静电能。由于典型的物体根据大小排列靠近,所有的能量都汇集到一起。水平线显示了热能大小以KT计,当然并不依据物体大小规模。为方便比较,图中显示了氢结合下的有规则的结合能,三磷酸腺苷中的磷酸盐组,共原子结合(环),还有典型的原子和亚原子能。
在分子机器与肉眼所见的对应物之间最重要的区别之一,是分子机器生活在一个热能的环境中。作为热能与确定性能之间相互影响的结果,统计力学是理解分子机器的必需工具。为了理解热效应的重要性,请记住物质的生物学能量单元是微微牛顿/毫微米(pN/nm),其中微微牛顿是分子机器所产生的特有能量,最终通过单细胞试验加以确定,而毫微米是典型的长度刻度。分子机器以100%的效率运行,每个循环使用一个ATP,产生约100 pN/nm的功。相比较而言,热能单位KT约为4个pN/nm。
进化的成功之一就是大自然发现了如何在非常“嘈杂”的环境中制造出高精确度的机器。一个不可思议的例证就是DNA聚合酶,一台仅有13毫微米大小的分子复制机器,能够以近百万分之一的误差复制DNA分子。这种令人叹为观止的保真度,特别是在试管或者细胞内的热环境下做到这一点的。当然还有很多的问题有待弄清。
分子机器能够在噪音中保持精确运作,而且借助波动也是它们的基本功能之一。例如限制酶,一种能识别并切除特定DNA序列的蛋白质。那些酶能搜寻由数以百万计、甚至数十亿核苷酸组成的基因组,并在与它们的识别序列结合时特别有效,而且完成任务的速度与顺着DNA分子的一维散射或三维散射并结合到DNA的目标序列不相匹配。取而代之的是,限制酶利用导致长DNA分子绕成团的熵能,从一条链跳到另一条链,加速与一维扩散有关的搜索过程。
完整的分子机器的理论需要考虑图2所示的所有反应,必须包括连续力、统计力、化学动力学和液体力学的理论,进而提出预见性的、统一的框架系统,而不是求助于完整的原子描述,应允许对任何分子机器的动力学行为进行精确的描述。事实上对物理学家而言,研究分子机器最迷人的挑战之一就是在何种程度上首先寻找到能构建分子机器的粗糙模型。
不是由单个细胞构成的
在理解分子机器在细胞内运行的过程中,科学家确认了许多有利于ATP活动和产生力量-速率的结构(有特色的单个发动机),还探究了转化和功能之间的联系。但理解单个机器和机器集合后是如何增添生物体活动的问题上,虽然许多定量模型视细胞为“酶包”,事实上细胞有着丰富的内部结构。
生物学功能的主要特点之一是在空间和时间上的排序,至少两类生物学编组能呼唤物理学家行动起来:物理结构和过程的协同以及信息的编组。在某种意义上,我们提供了描述生命机器时的对应物。在讨论中我们赞美了分子机器以及科学家必须面对的挑战。形成鲜明对比的是,有人争辩道,即使对每个细胞机器有正确的理解,仍然不足以解释细胞内部究竟发生了什么,就如同理解氢原子仅仅是解释结晶体内电子行为开始的序曲。引人注目的是,分子机器的集体效应如同量子行为的霍尔效应。
在许多例子里,细胞机器是许多部件的整合,它们经常是蛋白质、核酸,脂质和其他的协同物质的分子。物理学家了解这些分子相互作用的最重要的方式之一是考虑集体激发现象。如结晶体振动的声子特色和磁振子描述磁纤维的集体激发等。
实际上,物理学家谈到的各种各样的“-ons”,生物学装置也提供了类似的对应物。因为某些生物学结构有“-体”的标签,指的是源于多分子成分的大分子集合,大分子集合以集体形式执行多种功能。值得注意的是用于蛋白质合成中的核糖体;包裹真核生物DNA的最小单元——核小体;调节蛋白质降解装置的蛋白酶体和调节基因转录的转录体。通过目前还不为人知的机制和原理,蛋白质装配成“-体”,执行任务,然后再次分解。
最令人兴奋的是生物学协同行动的中心法则被揭示,即核糖核酸的复制和转录成信使核糖核酸(转录),最后由核糖体把信使核糖核酸转成相应的蛋白质(翻译过程)这一整套过程。这一过程包括不同层次的协同工作,从大分子联合体的装配到机器群的同时作用,再到这一过程中细胞可能采取的协作方式。图3显示了处于转录和翻译过程中遵循中心法则的(细菌)机器群。
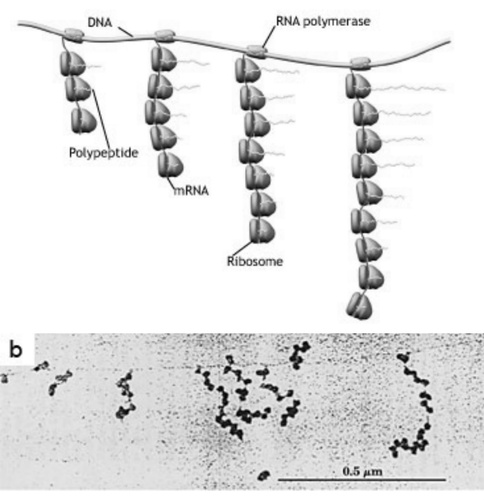
图3. 协同作用中大肠埃希菌中的分子机器。在草图中,转录过程中RNA聚合酶移动到DNA右侧并且缠绕在信使RNA上。同时,分子机器参与翻译:核糖体沿着RNA移动并且制造多肽。最后,那些多肽将组成蛋白质。(b)转录和翻译同时发生的电子显微镜照片。
协同行动的主题在生物系统的信息流中也得到了披露。如卵子分裂成胚胎这一在信号传导中处理的信息,在空间和时间上的要求是十分精确的。生物学信号的传导常常通过一组组动画呈现出来,各种各样的蛋白质信号通过不太被人理解的方式相互联系着,其表现形式:先是点对点附着进而组成网络。尽管对其所知甚少,我们应该有可能开创理解这一过程的理论。事实上,对生物学网络的总体分析——系统生物学——正使得生物学界为之兴奋。
中心法则里的信息流同样以动画的形式展现出来:一系列定向的箭头显示信息从DNA到RNA再到蛋白质,和从DNA到DNA。但是信息流也从蛋白质流向DNA,因为蛋白质通过各种结合DNA的方式调节基因的表达。虽然生物学家知道信息流的有趣特征,但中心法则的画面省略了封闭环的箭头。这种省略对理论的形成至关重要的。
而要理解细胞中的协同效应应整合两种哲学观点:第一,生命如同计算机程序,是机器中执行指令的部分,这些指令被编码成DNA软件。第二,纯物理学的观点,生命起源于化学物质的混合,这种混合遵循了基本的物理学原则。据推测,后者可利用的行为指令系统很有限。这两种观点是互补的,在特定的场合下,其中的一种可能比另一种能更好的解释细胞的行为。
空间中的时间
一种流行的捕获细胞机器的生物学思考方法,是研究细胞结构与功能之间的联系。随着越来越多的发现,结构问题得到了解决;而功能是关于动力学的固有问题,目前这一问题基本上尚未得到解答。因此对那些关注细胞机器功能的人而言,最富挑战的莫过于将时间引入恰当的地方。更深层次讲,大分子的动力学、它们之间的组合、细胞机器以及细胞本身的难题呈现在物理学家面前,要求他们在更大程度上解释这个看似不平衡的系统。事实上,我们相信生物动力学在物理学中是一个不平衡的案例。直到目前,在不平衡系统的研究中,重点仍放在违背平衡的问题上。此外,围绕不平衡问题的争论比预测特定的实验更复杂。尽管如此,生物学仍然可以提供有关不平衡物理学的系统性和预测性的思考起点。
欧文 · 薛定谔(Erwin Schrdinger)赞同要想理解生物学就要理解不平衡系统,他在发表于1944年的经典论文《生命是什么?》(剑桥大学出版社,1992)中阐述了这一观点。他认为新的物理学理论应关注对单一分子行为的理解,而远不是平衡问题。当薛定谔写这篇论文时,科学家们还不知道组成生命基础的分子特性。直到现在,人们可以推断基因是一种分子,并且认识到理解生命的机制有赖于对分子机器的认识。几种思考方法也许可以应用到不平衡系统的主题上。悲观的观点认为对普遍原则的研究注定会失败,没有什么比解决一个一个案例更好。有观察人士对这一观点不以为然。例如,物理学家珀西 · 布里奇曼(Percy Bridgman)论述道:“在不可逆转的进程中普遍的无能是一件令人吃惊的事情,物理学一般不接受这种失败主义态度。当然这也许是一种说词,如果他选择这样,那么依据定义人们可以说热力学仅仅在研究平衡状态。但是这种措辞毫无用处,物理学不会因为研究不可逆转进程而被抹去。”由于生物系统本质上是不平衡的,故生物系统的研究就要求物理学家加倍努力在不平衡进程的研究中取得进展。当然,对不平衡行为的生物学案例进行定量分析的机会将越来越多。
细胞内部的重要特征也使得细胞学研究充满挑战性,这一点在大卫 · 古德塞尔(David Goodsell)的画作中得到完美地描绘。如平衡态统计力学的标准仪器需要解决可移动细胞前缘的动态组合体。它们不但很不平衡,而且适合该系统的标准方法经常受控于以稀溶液和扩散方程为基础的化学能。结果,对活细胞内部既拥挤又忙乱的研究产生了大量关于物理材料的疑问,这些物理材料既不稀松,也不静态,更不均一。随着越来越富有规律性,对单一分子、大分子组合,甚至细胞本身的实验观察都按照原有轨道有所前倾。也就是说,科学家已经认识到,生物系统的暂时演变是有趣的、可以测量的和可再生的。如最近的一组实验(图4左)显示,对单一分子发动机轨道的长期监测是可能的,并且能获得非常高的空间分辨率。这类实验不仅取得了发动机运转的机制,还对发动机的协同作用以及单一发动机经历的波动现象有所揭露。图4右证明了轨道分析在较大范围内仍然有用。事实上,可移动细胞的轨道暴露在随时间变化的温度下,而且提供了有关骨架蛋白行为的动力学和控制的线索。
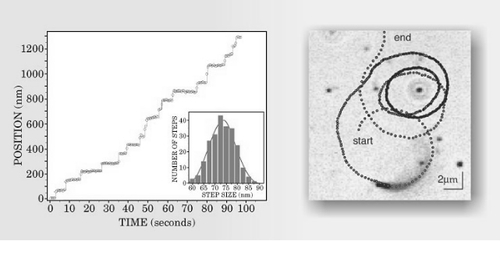
图4. 生物体轨道能被精确测量。左图为单一分子发动机阻凝蛋白作为计时功能时的位置,能通过检测依附于发动机的分子荧光强度加以确定。右图为等时步骤在利斯特菌轨道中得到显示和细菌把主体细胞构建成肌动蛋白网络。
动画以外
自狄拉克引发挑战以来的75年来,科学家已经在发现和形成生命基础的分子编码方面取得了巨大的进展。他们的富有智慧的追寻与前分子时期的“集邮”有何不同?生物学信息大爆炸提供的一个最大机会就是再探生物学现象并在理论和实验中使用定量互动的方法以提高理解。本文列举了物理学家能服从于实验和理论的领域包括:理解分子机器和分子组合的运转原则、理解细胞一生引发的空间和时间完美组合的协同效应,以及产生有关不平衡状态统计机制的观念和想法,这些观念和想法为理解体细胞的变化提供了适宜的框架。很明显,生物学家已经在那些议题上有了较深入的思考,但是我们相信,从物理学角度来思考生命现象,能给自身带来独特的贡献。
众多实验技术的涌现为生物体的研究打开了一扇窗户。完整的基因组测序和基因表达分析暗示生物体是一个有限的、封闭的系统,而且相关数据也限制了可能的模型数。此外,人们能够详尽地测量细胞以及与环境混乱引发的后果。这种现象接受一种新的理论观点:对混乱的后果进行系统分析而非不完整的、割裂的评估。单一分子的生物-物理学技术,产生了新的观察和学习方法,并且为大分子机器增添了特色,为解决在本文中提及的一些问题提供了数据。我们确信,这些问题将在相当长的时间里向实验和理论物理学家发出猛烈的挑战。
[作者介绍:罗布·菲利普斯(Rob Phillips)是加州理工学院的应用物理学和机械工程学教授。史蒂夫·奎克(Steve Quake)是斯坦福大学应用物理系生物工程教授、霍华德-休斯医学研究中心的研究员。]